Global Fisheries and the Growth of Greenhouse Gas Emissions
By Samantha Orndorff, SRC intern
Global fisheries have long been fundamental in molding cultural identities, maintaining economic sustainability, and providing a reliable source for food production. The distribution and production of food, such as that generated from fisheries, is responsible for a quarter of anthropogenic greenhouse gas (GHG) emissions (Parker et. al 2018). As climate change issues become more paramount, it is imperative that systems of emission are studied to develop management strategies and initiatives to mitigate environmental impact. Fisheries is typically an energy-intensive operation that produces the majority of its emissions directly from burning fossil fuels. A recent study conducted by the Institute for Marine and Antarctic Studies analyzed fuel use data from a Fisheries Energy Use Database in order to quantify fuel input and greenhouse gas emissions produced by the global fishing fleet from 1990-2011. Despite the fact that harvest has remained relatively stable over the past two decades, researchers have found that GHG emissions from world fisheries has increased by 28% from 1990 to 2011.
The countries with the largest national fishing fleets are China, Indonesia, Vietnam, the United States, and Japan. In 2011 approximately 49% of total fishery GHG emissions came solely from the contributions of the five aforementioned countries (Figure 1). Emissions by fishing sector vary considerably based upon targeted species class. For example, crustacean fisheries such as those targeting lobster and shrimp harvest a smaller volume than that of a small pelagic fisheries targeting menhaden which are easily caught. Crustacean fisheries tend to account for a larger percentage of fishery GHG emissions because of the considerable amount of fuel required to target such high-input species. In fact, much of the overall increase in emissions from 1990 to 2011 can be attributed to changes in catch composition, with crustacean catch rates increasing by 60% over two decades (Parker et. al 2018). Thus, the GHG emissions from Asian fishing fleets are much more substantial than European and American fishing fleets given that Asian countries disproportionately target crustaceans whereas Europe and the Americas are primarily comprised of low-input small pelagic fisheries.
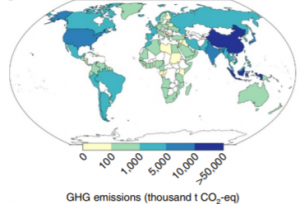
Figure 1. GHG emissions in 2011 for each national fishing fleet, up to the point of landing in thousands of tons of carbon dioxide (thousand t CO2-eq) (Parker et. al 2018).
When analyzing the global GHG emissions from other sources of animal protein, such as that of pork, beef and lamb, products derived from marine fisheries for human consumption have significantly lower GHG emissions (Figure 2). It is hypothesized that if fish landed for non-food consumption, such as those used in meal and oil production for aquaculture and livestock, were directed for human consumption than total fisheries emissions would be “lower than every other major source of animal protein” (Parker et. al 2018). Furthermore, global emissions from agriculture and livestock production amounted to 5 billion tons of carbon dioxide in 2011 whereas emissions from marine fisheries only amounted to 179 million tons of carbon dioxide.
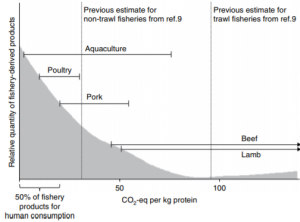
Figure 2. Carbon footprint of fishery-derived products for human consumption in 2011 compared to other sources of animal protein (Parker et. al 2018).
Proposed strategies to mitigate GHG emissions from global fisheries include rebuilding fishery stocks and reducing quotas so that the amount of fuel utilized by national fleets can be reduced. Short-term adaptations to improve emission reductions are using more selective fishing times and locations to optimize landings and fuel use. Further and continued research will also aid in creating a more long-term, dynamic and comprehensive solution for studying the relationship between global fisheries and total greenhouse gas emissions.
Work Cited
Robert W. R. Parker, Julia L. Blanchard, Caleb Gardner, Bridget S. Green, Klaas Hartmann, Peter H. Tyedmers & Reg A. Watson. April 2018. Fuel use and greenhouse gas emissions of world fisheries. Nature Climate Change 8:333-337.