Effect of Climate Change on Pacific Tuna Stocks
By Beau Marsh, RJD Intern
Pacific tuna fisheries are essential to the livelihood, sustainability and well-being of many Pacific islands, they acts as an important food source, as well as a lucrative activity for local economies. The species of tuna composing these fisheries are Skipjack (Katsuwonus pelamis), Yellowfin (Thunnus albacares), Bigeye (Thunnus obesus), and Albacore (Thunnus alalunga). Skipjack are particularly significant because they regularly make up more than 60% of the Pacific tuna catch (Ganachaud et al., 2013). It is important to learn as much as possible about the conditions in which the different tuna species spend their time, so habitat shifts in response to climate change and El Nino Southern Oscillation (ENSO) can be anticipated.
Tuna are considered thermoregulators (Lehodey et al., 2010). This means that they can tolerate a greater range of temperatures by maintaining a fairly constant internal temperature. The mechanism by which this is possible is called countercurrent heat exchange, through which their circulatory system conserves heat more effectively (Lehodey et al, 2010). This mechanism is more developed in certain tuna species, so different species occupy different parts of the water column based on temperature. Adult Skipjack reside in waters ranging from 20-29 degrees Celsius (Ganachaud et al., 2013). Other species, including Bigeye and Albacore, are capable of living at cooler temperatures, allowing them to dive to deeper depths. Bigeyes can dive to around 600 meters in about 5 degree Celsius water (Brill et al., 2005), whereas Skipjack are restricted to the upper 200 meters (Lehodey et al., 2010).
These environmental conditions do not apply to tuna larvae. In the larval stage, tuna are more sensitive to environmental factors. They require stricter physical and chemical conditions. The different species’ larvae spend all their time at the surface where the warmest water and food supplies are consistent (Brill et al, 2005). Distributions of tuna depend on horizontal stratifications by temperature, as well as the depth of the mixed layer (Ganachaud et al., 2013). Ideal conditions are formed by the convergence and divergence of ocean currents that create thermal fronts, locations of upwelling, and eddies (Langley et al, 2009). These physical boundaries, combined with foraging areas, dictate tuna distributions basin-wide.
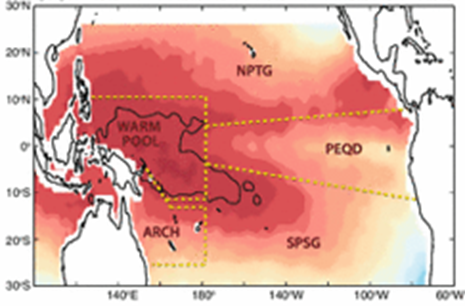
The physical boundaries are the Pacific Equatorial Divergence (PEQD), the West Pacific Warm Pool (WPWP), the North Pacific Subtropical Gyre (NPSG), and the South Pacific Subtropical Gyre (SPSG) (Ganachaud et al., 2013). In the case of the Skipjack, over 90% of the population resides in the WPWP, specifically, along the eastern front at its boundary with the PEQD (Fig.1) (Ganachaud et al., 2013).
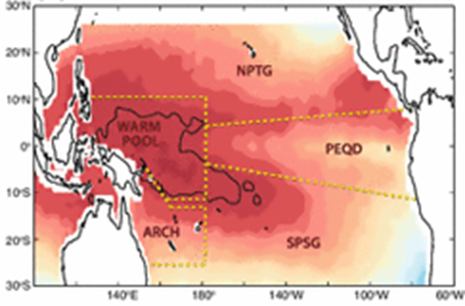
SST and superimposed Pacific hydrological factors (Ganachaud et al, 2013).
In contrast to the WPWP, the waters of the central and eastern Pacific are characterized by higher salinity (>35 psu) and increased concentration of nutrients (Maes et al, 2006). This is explained by the strong wind-driven upwelling that takes place along the west coast of South America and the Ekman-driven divergence that occurs in the Central Equatorial Pacific (Picaut et al, 2001). Due to the strong difference between the WPWP and the PEQD, the boundary of these water masses exhibits unique physical, chemical and biological characteristics and is the location of most tuna fisheries in the Pacific (Lehodey et al, 2010). Although the abundance of tuna in warm, oligotrophic waters in the WPWP is counter-intuitive at first glance, convergence zones such as the one between the WPWP and the PEQD are known to act as aggregating mechanisms of plankton and micronekton and, subsequently, large predators.The western equatorial Pacific presents one of the most prolific areas for tuna in terms of spawning and foraging grounds (Ganachaud et al, 2013). Formed by the wind-driven South Equatorial Current (SEC), the WPWP is characterized by the highest sea surface temperature of the world’s oceans (often reaching more than 30°C) and low sea surface salinity caused by increased precipitation (McPhaden and Picaut, 1990; Picaut et al, 2001; Maes et al, 2006). As a result of this accumulation of water in the western Pacific, the thermocline in the WPWP is considerably deeper than along the central and eastern Pacific; this inhibits deep vertical mixing and makes the WPWP an area of low nutrients and low primary productivity (Picaut et al, 2001).
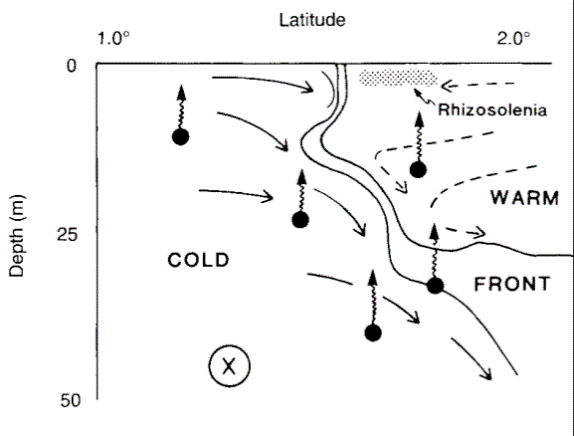
Schematic drawing illustrating a possible aggregation mechanism for plankton and micronekton (Yoder et al, 1994)
The WPWP-PEQD convergence boundary is not static; it’s modulated by the extent and location of the WPWP and has a well defined interannual variability that is strongly linked to the phase of ENSO. McPhaden and Picaut (1990) first observed the migration of the WPWP in relation to ENSO; they reported a strong eastward displacement of the eastern boundary of the WPWP during El Niño and associated this shift to a reversal of the SEC caused by weakened trade winds. Later, Lehodey et al (2010) reported a strong correlation between the location of the WPWP-PEQD boundary (using the 29° C isotherm as a proxy), the abundance of Skipjack tuna and the phase of ENSO; they reported clear eastward population shifts during the warm phase of ENSO. Other approaches have been used to ascertain the location of the convergence zone and its variability; Maes et al (2006) identified a strong salinity gradient (0.4 psu over 10°-15° in longitude) to be a more accurate indicator, and more recently have used satellite-based ocean color observations to detect the eastern edge of the WPWP (Maes et al, 2006).
In an attempt to incorporate climate change predictions into tuna population models, Lehodey et al (2010) forecast the distribution of Bigeye tuna in the Pacific. Results of this study showed an expansion of favorable spawning area towards mid-latitudes and to the eastern Pacific; in contrast, the WPWP became too warm and oxygen depleted to sustain stable populations of tuna by the end of the century.
Ganachaud et al (2013) documented changes in oceanographic parameters of importance to tuna distribution in the Pacific. They reported a large-scale shoaling of the thermocline, increasing stratification and limiting nutrient provision to the biologically active layer. They also highlight changes in oceanic currents such as the strengthening of the NEC which they hypothesize could modify the supply of iron in the eastern Pacific. Ganachaud et al (2013) anticipate an eastward migration of the WPWP-PEQD boundary by 6,000 (±2,000) km.
Global warming will affect the ocean’s physical, chemical and biological characteristics and those changes will have an impact on fisheries and its fish-dependent communities (Hollowed at al, 2013). The extent to which fisheries will be able to continuously provide food for millions of people is contingent upon our understanding of future distributions and abundance of commercially important species such as tuna.
Attempts should be made to provide new insight into future spawning and foraging favorable grounds for tuna in the Pacific Ocean. Traditionally, the areas near the NPSG and the SPSG have been overlooked and categorized as low productivity areas. Nevertheless, the NPSG-PEQD and SPSG-PEQD boundaries might exhibit similar oceanographic characteristics to those found in the convergence zone at the WPWP-PEQD boundary and it is possible that under a global warming scenario these areas will become more favorable for tuna populations. Efforts should be made to ascertain the suitability of these regions as potential future tuna habitats.
References
Brill, R. W., Bigelow, K. A., Musyl, M. K., Fritsches, K. A., & Warrant, E. J. (2005). Bigeye Tuna (Thunnus obesus) Behavior And Physiology And Their Relevance To Stock Assessments And Fishery Biology, 57(2), 142–161.
Ganachaud, A., Sen Gupta, A., Brown, J., Evans, K., Maes, C., Muir, L. C., & Graham, F. S. (2013). Projected changes in the tropical Pacific Ocean of importance to tune fisheries. Climatic Change, 119, 163-179.
Hollowed, A., Barange, M., Beamish, R., Brander, K., Cochrane, K., Drinkwater, K., Foreman,G.,Hare,J., Holt,J., Ito,S., Kim,S., King,J.R.,Loeng,H., MacKenzie,B.R., Mueter,F.J., Okey,T.A., Peck, M., Radchencko,V.I., Rice,J.C., Schirripa,M.J., Yatsu,Y., Yamanaka,Y.. (2013). Projected impacts of climate change on marine fish and fisheries. ICES Journal of Marine Science, 70(5), 1023-1037.
Langley, A., Briand, K., Sean, K. D., & Murtugudde, R. (2009). Influence of oceanographic variability on recruitment of yellowfin tuna (thunnus albacares) in the western and central Pacific Ocean. Canadian Journal of Fisheries and Aquatic Sciences, 66, 1462-1477.
Lehodey, P., Senina, I., Sibert, J., Bopp, L., Calmettes, B., Hampton, J., & Murtugudde, R. (2010). Preliminary forecasts of bigeye tuna population trends under the A2 IPCC scenario. Progress in oceanography, 86, 302-315.
Maes, C., Ando, K., Delcroix, T., Kessler, W., McPhaden, M., & Roemmich, D. (2006). Observed correlation of surface salinity, temperature and barrier layer at the eastern edge of the western Pacific warm pool. Geophysical Research Letters, 33. doi: 10.1020/2005GL024772
McPhaden, M., & Picaut, J. (1990). El Nino-Southern Oscillation Displacements of the Western Equatorial Pacific Warm Pool. Science, 250, 1385-1388.
Picaut, J., Ioualalen, M., Delcroix, T., Masia, F., Murtugudde, R., & Vialar, J. (2001). The oceanic zone of convergence on the eastern edge of the Pacific warm pool: A synthesis of results and implications for El Nino-Southern Oscillations and biogeochemical phenomena. Journal of Geophysical Research., 106, 2363-2386.
Yoder, J., Ackleson, S., Barber, R., Flament, P., & Balch, W. (1994). A line in the sea. Nature, 371, 689-692.


Leave a Reply
Want to join the discussion?Feel free to contribute!