
Re-evaluating the health of coral reef communities: baselines and evidence for human impacts across the central Pacific
By Shannon Moorhead, SRC Masters Student
In the past several decades, it has become clear to researchers that populations of reef-building corals have suffered significant declines worldwide. In the 1970s, coral covered on average,50% of benthic habitat (the sea floor) in the Caribbean; in the early 2000s, this was reduced to an average of 10%, with an estimated 80% decline in total cover throughout the Caribbean. Similar observations have been made in the Pacific, with an estimated decrease from 43% coral cover on average in the 1980s, to 22% cover on average in 2003. These declines are caused by a large variety of both global and local threats. Globally, increasing temperatures, ocean acidification, sea level rise, and disease outbreaks have caused mass coral mortality events and decreased rates of calcification – the rate at which coral grows its calcium carbonate skeleton. In addition to these worldwide stressors, corals face local threats such as overfishing of important grazers and predators, elevated nutrient levels, and increasing amounts of terrestrial sediment in coastal waters. These human impacts kill corals either directly or indirectly, by creating conditions that allow faster-growing algae species to thrive and overtake corals. In some cases, this algal growth can lead to a phase-shift: a change in ecosystem composition and function when macroalgae replaces corals as the dominant benthic cover. Because of this, the majority of studies done to assess reef health have focused only on percent cover of macroalgae and corals. However, recent research indicates that when coral cover declines it is rarely replaced solely by macroalgae and studies have shown that coral and macroalgae together only comprise 19-55% of the reef benthos, organisms that live on the sea floor.

Figure 1. (a) Theses images show hard coral and macroalgae, the two groups most often used to assess reef health. (b) These images show reef builders and fleshy algae, which can be used to assess reef-health with a more community-based approach.
Methods
In this study, Smith et al. also investigated percent cover of other members of the benthos: crustose coralline algae (CCA) and turf algae. Turf algae have a negative impact on coral cover, by growing over and smothering adult corals and preventing the settlement of larval corals. On the other hand, CCA, which produces calcium carbonate like corals, promotes reef resilience by stabilizing reef structure and creating a place for coral larvae to reside and grow, because there are many coral species whose larvae prefer to settle on some types of CCA. Smith et al. considered the cover of all four groups (coral, CCA, turf algae, and macroalgae) to compare central Pacific reef communities surrounding uninhabited islands with communities that surround populated islands and suffer from significant anthropogenic, or human-caused, stressors. Specifically, they examined whether cover of reef-builders (CCA and coral) and fleshy algae (turf and macroalgae) were inversely related, as well as whether the two groups were more common in the absence or presence of human populations.
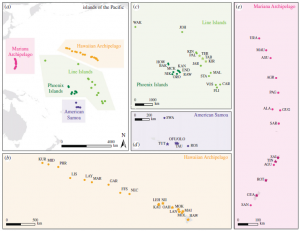
Figure 2. (a) A map of the five island chains and 56 islands from which data were collected for this study; (b) 17 Hawaiian Islands, (c) 21 islands from the Line and Phoenix Islands, (d) six islands from American Samoa, and (e) 14 islands in the Mariana Archipelago. Stars represent inhabited islands while circles represent uninhabited islands.
Results
Smith et al. evaluated reef communities of 56 islands from five central Pacific island chains between 2002 and 2009 and acquired some surprising results. While coral cover was higher on uninhabited islands, there was not a significant difference between the two. Macroalgae cover varied by archipelago: while there was greater macroalgae cover on populated islands in the Line and Mariana Islands, the Hawaiian Islands and American Samoa had higher macroalgae cover on uninhabited islands. In addition, there was no significant relation seen between total coral and macroalgae cover, contradicting previous ideas that macroalgae directly replaced coral on degraded reefs. However, the average cover of reef-builders was significantly higher on uninhabited islands versus inhabited islands, while the average cover of fleshy algae was significantly higher on inhabited islands versus uninhabited. This result suggests that local anthropogenic stressors play a direct role in changes to the benthic community, and potentially reef health.
Outcomes
In this study, the authors suggest that a good indicator of reef health is net accretion, where the reef-building organisms are building calcium carbonate skeletons faster than they are being eroded. Because it appears that inhabited islands have a lower abundance of reef-building organisms, the reefs surrounding these islands are not as healthy as those near uninhabited islands and may have a harder time bouncing back from large-scale disturbances such as bleaching events and typhoons. Local management on inhabited islands should consider this when developing management strategies and work towards improving the resilience of their reef ecosystems. This research also demonstrates that percent coral and macroalgae cover are not always reliable indicators of reef health; instead management should take a more holistic approach and evaluate other members of the benthos when assessing reef health, in addition to measuring indicators of reef resilience such as coral growth and recruitment, which will help managers predict trends and changes in the structure of the benthic community of coral reefs.
References
Smith, J. E., Brainard, R., Carter, A., Grillo, S., Edwards, C., Harris, J., . . . Sandin, S. (2016). Re-evaluating the health of coral reef communities: baselines and evidence for human impacts across the central Pacific. Proceedings of the Royal Society of London B.


Leave a Reply
Want to join the discussion?Feel free to contribute!